El
agua
La
vida en la Tierra se suele describir como un fenómeno basado en el carbono, pero
sería igualmente correcto llamarlo un fenómeno basado en el agua. Es probable que
la vida se haya originado en el agua, hace más de tres mil millones de años, y
que todas las células vivientes sigan dependiendo del agua para existir. En la mayor
parte de las células el agua es la molécula más importante y forma de 60 a 90% de
su masa, aunque hay pocas excepciones, como las semillas y las esporas, de las
cuales se expulsa el agua. Las semillas y las esporas pueden permanecer
latentes por largos periodos hasta “revivir” por la reintroducción de agua.

·
La
molécula de agua es polar
La molécula de agua (H2O) tiene
forma de V y el ángulo entre los dos enlaces covalentes O—H es de 104.5°.
Algunas propiedades importantes del agua se deben a la forma angulada y a los
enlaces intermoleculares que puede formar. Un átomo de oxígeno tiene ocho
electrones y su núcleo cuenta con ocho protones y ocho neutrones.
Hay dos electrones en la capa
interna y seis en la capa externa de este átomo. La capa externa presenta el
potencial de contener cuatro pares de electrones en un orbital s y tres
orbitales p. Sin embargo, la estructura y las propiedades del agua se
pueden explicar mejor si se supone que los electrones de la capa externa ocupan
cuatro orbitales híbridos sp3.
La polaridad de una molécula
depende tanto de la polaridad de sus enlaces covalentes como de su geometría.
La disposición angulada de los enlaces polares O—H del agua produce un dipolo
permanente para toda la molécula.
Una molécula de amoniaco también
contiene un dipolo permanente. Así, aun
cuando el agua y el amoniaco gaseoso son eléctricamente neutros, ambas
moléculas son polares. La gran solubilidad de las moléculas polares de amoniaco
en el agua se ve facilitada por fuertes interacciones con las moléculas polares
del agua.
·
Puentes
de hidrógeno en el agua
Una de las consecuencias
importantes de la polaridad de la molécula de agua es que dichas moléculas se
atraen entre sí. La atracción entre uno de los átomos de hidrógeno, ligeramente
positivo, de una molécula de agua y los pares de electrones parcialmente negativos
en uno de los orbitales híbridos sp3, produce un “puente de hidrógeno”. En
un puente de hidrógeno entre dos moléculas de agua, el átomo de
hidrógeno permanece enlazado covalentemente a su átomo de oxígeno que es el
donador de hidrógeno. Al mismo tiempo, está unido a otro átomo de oxígeno,
llamado aceptor de hidrógeno. De hecho, el átomo de hidrógeno está compartido
(en forma desigual) entre los dos átomos de oxígeno.
El agua no es la única molécula
capaz de formar puentes de hidrógeno; esas interacciones pueden existir entre
cualquier átomo electronegativo y un átomo de hidrógeno unido a otro átomo
electronegativo. (Se examinan otros ejemplos de puentes de hidrógeno en la
sección 2.5B). Los puentes de hidrógeno son mucho más débiles que los enlaces
covalentes típicos.
En los puentes de hidrógeno, la
orientación es importante. Un puente de hidrógeno es más estable cuando el
átomo de hidrógeno y los dos átomos electronegativos asociados a él (los dos
átomos de oxígeno, en el caso del agua) forman casi una línea recta. Una sola molécula
de agua puede formar puentes de hidrógeno hasta con otras cuatro moléculas de
agua.
La capacidad que tienen las
moléculas de agua en el hielo para formar cuatro puentes de hidrógeno y la fuerza
de éstos determinan que el hielo presente un punto de fusión anormalmente alto.
Ello se debe a la gran cantidad de energía que se requiere en forma de calor
para destruir la red cristalina del hielo a base de puentes de hidrógeno.


·
El
agua es un solvente excelente
Las propiedades físicas del agua
se combinan para hacerla un excelente solvente. Ya se estudió que las moléculas
de agua son polares; esta propiedad ejerce consecuencias importantes, como se
verá después.
A. Las
sustancias iónicas y polares se disuelven en agua
El agua puede interactuar y
disolver otros compuestos polares y compuestos que se ionizan.
La ionización se relaciona con la
ganancia o pérdida de un electrón, que da lugar a un átomo o a un compuesto que
presenta una carga neta. Las moléculas que se pueden disociar y formar iones se
llaman electrólitos. Las sustancias que se disuelven con facilidad en
agua se llaman hidrofílicas o amantes del agua.
Los electrólitos no son las
únicas sustancias hidrofílicas que son solubles en agua.
Toda molécula polar también
exhibirá una tendencia a solvatarse por moléculas de agua. Además, la
solubilidad de muchas moléculas orgánicas aumenta por la formación de puentes
de hidrógeno con las moléculas de agua.
La cantidad de grupos polares en
una molécula afecta su solubilidad en agua. También la solubilidad depende de
la relación entre grupos polares y no polares en una molécula.
B. Concentraciones
celulares y difusión
El comportamiento de los solutos
en el citoplasma es distinto del que tienen en una sencilla solución en agua.
Una de las diferencias más importantes es la reducción de la velocidad de
difusión dentro de las células.
Hay tres razones por las que los
solutos se disuelven con más lentitud en las células.
1. La viscosidad
del citoplasma es mayor que la del agua, lo que se debe a la presencia de
numerosos solutos, como los azúcares. De acuerdo con mediciones recientes
parece que la viscosidad del citoplasma sólo es un poco mayor que la del agua,
aun en los organelos empacados densamente.
2. Las moléculas
con carga se enlazan momentáneamente entre sí dentro de las células y ello
restringe su movilidad. Dichas consecuencias de la unión ejercen un efecto
pequeño, pero apreciable, sobre las tasas de difusión.
3. Los choques con
moléculas de agua inhiben la difusión a causa de un efecto que se denomina hacinamiento
molecular. Es la principal razón por la que se desacelera la difusión en el
citoplasma.
C. Presión
osmótica
Si una membrana permeable al
solvente separa a dos soluciones que contienen concentraciones distintas de
sustancias disueltas, o solutos, las moléculas del solvente se difundirán desde
la solución menos concentrada hacia la más concentrada en un proceso llamado ósmosis.
La presión necesaria para evitar este flujo de solvente se llama presión
osmótica. La presión osmótica de una solución depende de la
concentración molar total del soluto y no de su naturaleza química.
Las sustancias no polares son
insolubles en agua
Los hidrocarburos y otras
sustancias no polares presentan una solubilidad muy baja en agua porque las
moléculas de agua tienden a interactuar con otras moléculas de agua y no con
moléculas no polares. El resultado es que las moléculas de agua excluyen a las sustancias
no polares forzándolas a asociarse entre sí. Por ejemplo, las gotas diminutas de
aceite que se dispersan en forma vigorosa en agua tienden a coalescer y formar
una sola gota, con lo cual minimizan la superficie de contacto entre las dos
sustancias. Es la explicación por la que se separa el aceite en un aderezo para
ensalada si se deja en reposo durante algún tiempo antes de agregarlo en la
ensalada.
La acción limpiadora de los jabones
y demás detergentes se deriva de su capacidad de atrapar la grasa y los
aceites, insolubles en agua, en los interiores hidrofóbicos de las micelas. El
SDS y los detergentes sintéticos similares son ingredientes activos comunes en
detergentes para lavandería. La suspensión de compuestos no polares en agua,
por su incorporación a las micelas, se llama solubilización, que no es lo mismo
que disolución. Varias estructuras que se encontrarán después en este libro,
como proteínas y membranas biológicas, se asemejan a las micelas por exhibir
interiores hidrofóbicos y superficies
hidrofílicas.
PROTEÍNAS
Las proteínas son macromoléculas compuestas
por carbono, hidrógeno, oxígeno y nitrógeno.
Las mismas están formadas por la
unión de varios aminoácidos, unidos mediante enlaces peptídicos.
El orden y disposición de los aminoácidos en una proteína depend del código genético,
ADN, de la persona.
Las proteínas son las moléculas
biológicas más abundantes y tienen diversas formas, tamaño y funciones.
Las enzimas son las más variadas
y especializadas. Prácticamente todas las reacciones celulares están
catalizadas por proteínas.
Una proteína puede ser una sola
cadena polipeptídica o puede estar formada por varias de esas cadenas unidas
entre sí por interacciones débiles. Como regla general, cada cadena
polipeptídica es codificada por un solo gen, aunque hay algunas excepciones interesantes
a esta regla. El tamaño de los genes y los polipéptidos que codifican puede variar
en más de un orden de magnitud. Algunos polipéptidos sólo contienen 100
residuos de aminoácido, con una masa molecular relativa aproximada de 11 000 (Mr
_ 11
000). (La masa molecular relativa
de un residuo de aminoácido en una proteína es 110). Por otra parte, algunas
cadenas muy grandes de polipéptido contienen más de 2000 residuos de aminoácido
(Mr _ 220 000).
Las proteínas fibrosas son
una clase particular de proteínas estructurales que proporcionan soporte mecánico
a las células u organismos. En el caso típico, las proteínas fibrosas se
ensamblan en grandes cables o hebras. Como ejemplos de proteínas
fibrosas están la a-queratina, el componente principal de cabello y
uñas, y la colágena, el componente proteínico.
·
Hay
cuatro niveles de estructura de las proteínas
Las moléculas individuales de
proteína se pueden describir mediante hasta cuatro niveles de estructura:
La estructura tridimensional de
una proteína se describe con tres niveles adicionales: estructura secundaria,
estructura terciaria y estructura cuaternaria.
La estructura secundaria se
refiere a las regularidades en las conformaciones locales mantenidas por
puentes de hidrógeno entre los hidrógenos de amida y los oxígenos de carbonilo
en la columna vertebral del péptido. Las estructuras secundarias principales
son las hélices a y las hebras b (incluyendo las lámnas a). Se acostumbra
representar las regiones helicoidales a con dibujos que muestran las
estructuras de proteínas plegadas; las hebras b se representan con flechas
anchas que apuntan desde la dirección N-terminal hacia la C-terminal.
La estructura terciaria describe
la cadena polipeptídica totalmente plegada y compactada. Muchos polipéptidos
plegados consisten en varias unidades distintas unidas por un tramo corto
de residuos de aminoácidos, como se ve en la figura 4.1c; a dichas
unidades se les conoce como dominios. Las estructuras terciarias se
estabilizan por las interacciones de cadenas laterales de aminoácidos en
regiones no vecinas de la cadena polipeptídica. La formación de la estructura
terciaria acerca partes lejanas de las estructuras primaria y secundaria.
Algunas proteínas poseen estructura
cuaternaria, que implica la asociación de dos o más cadenas polipeptídica
en una multisubunidad, o proteína oligomérica u oligómera. Las cadenas polipeptídicas
de una proteína oligómera pueden ser idénticas o distintas.
Conformación del grupo peptídico
Nuestro estudio detallado de la
estructura de las proteínas comienza con la de los enlaces peptídicos, o enlaces de péptido, que unen a los aminoácidos
en una cadena polipeptídica. Los dos átomos que intervienen en el enlace peptídico,
junto con sus cuatro sustituyentes (el átomo de oxígeno carbonílico, el átomo
de hidrógeno de amida y los dos átomos adyacentes de carbono a) constituyen el
grupo peptídico. Los análisis cristalográficos de pequeños péptidos con rayos X
revelan que el enlace entre el carbono
carbonílico y el nitrógeno es más
corto que un enlace sencillo típico C—N, pero más largo que los dobles enlaces
C__N típicos. Además, el enlace entre el carbono carbonílico y el oxígeno es un
poco mayor que el doble enlace típico C__O. Esas mediciones indican que los
enlaces peptídicos tienen ciertas propiedades del enlace doble y se pueden representar
mejor como un híbrido de resonancia.


Aminoácidos
(aa)
·
Estructura
general de los aminoácidos
Todos los organismos emplean los
mismos 20 aminoácidos como bloques constructivos para armar las moléculas de
proteína. A estos 20 aminoácidos se les llama aminoácidos comunes, estándar
o normales. A pesar de la poca cantidad de los aminoácidos, se puede
obtener una variedad enorme de distintos polipéptidos al unir los 20
aminoácidos comunes para formar distintas combinaciones.
Los aminoácidos se llaman así
porque son derivados aminados de ácidos carboxílicos.
En los 20 aminoácidos comunes,
los grupos amino y carboxilo están unidos al mismo átomo de carbono: el átomo
de carbono alfa.
En 19 de los 20 aminoácidos que
se usan en la biosíntesis de proteínas, el átomo de carbono a es quiral,
o asimétrico, porque tiene cuatro grupos diferentes unidos a él. La excepción
es la glicina, cuyo grupo R sólo es un átomo de hidrógeno (la molécula no es quiral,
porque el átomo de carbono a está unida a dos átomos idénticos de hidrógeno). Los
19 aminoácidos quirales pueden, en consecuencia, existir como estereoisómeros.
·
Estructuras
de los 20 aminoácidos comunes
Las estructuras de los 20
aminoácidos que suelen encontrarse en las proteínas se muestran en las figuras
siguientes, como proyecciones de Fischer. En las proyecciones de
Fischer, los enlaces horizontales
en un centro quiral se extienden hacia el espectador y los verticales se
alejan. El examen de las estructuras revela que hay considerable variación en
las cadenas laterales de los 20 aminoácidos. Algunas cadenas laterales son no
polares, y en consecuencia hidrofóbicas, mientras que otras son polares o
ionizadas a pH neutro y en consecuencia son hidrofílicas. Las propiedades de
las cadenas laterales tienen gran influencia sobre la forma tridimensional general,
o conformación, de una proteína. Por ejemplo, la mayor parte de las cadenas laterales
hidrofóbicas de una proteína hidrosoluble se pliegan hacia el interior y le
proporcionan a la proteína una forma compacta y globular.
Es importante aprenderse las
estructuras de los aminoácidos estándar porque se mencionarán con frecuencia en
los capítulos sobre estructura de proteínas, enzimas y síntesis de proteínas.
En las secciones que siguen los aminoácidos estándar se agruparon por sus
propiedades generales y las estructuras químicas de sus cadenas laterales.
Las cadenas laterales pertenecen
a las siguientes clases químicas: alifática, aromática, sulfurada, alcohol,
base, ácido y amida. De los 20 aminoácidos, cinco se subdividen en muy (o altamente)
hidrofóbicos (en gris claro) y siete se clasifican como muy hidrofílicos (en
negro). Comprender la clasificación de los grupos R simplificará la
memorización de las estructuras y los nombres.
Aminoácidos
esenciales y No esenciales
Esenciales: histidina,
isoleucina, leucina, lisina, metionina, fenilalanina, treonina, triptófano y
valina.
No esenciales: alanina,
aspartato, cisteína, glutamato, glutamina, glicina, prolina, serina, tirosina,
asparagina
y arginina.

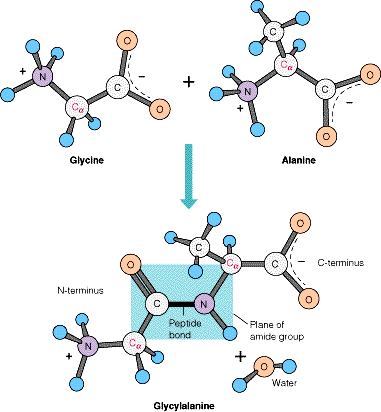
Péptidos
Los
péptidos son cadenas de aminoácidos
Ø El
enlace peptídico: Unión de dos aminoácidos
• Enlace covalente
tipo amida
• Reacción de
condensación
• Se elimina H2O
• La unión de dos aa
da lugar a un dipéptido
Ø El enlace
peptídico presenta un carácter parcial de doble enlace
El oxígeno del carbonilo tiene
una carga negativa parcial y el nitrógeno de la amida una carga positiva
parcial, la creación de un pequeño dipolo eléctrico. Prácticamente todos los enlaces
peptídicos de las proteínas se producen en esta configuración trans.
Existen
péptidos y polipéptidos biológicamente activos y de una gran variedad y tamaño.
Dipéptido sintetizado
comercialmente: Edulcorante artificial.


Enzimas
El nombre enzima deriva de una palabra griega que significa “en la levadura”. Indica que dichos catalizadores están presentes en el interior de las células. A finales del siglo XIX se estudió la fermentación de los azúcares por acción de células de levadura. Una generación después, James B. Summer cristalizó, en 1926, la primera enzima (ureasa) y demostró que era una proteína. En la siguiente década se purificaron cinco enzimas más y se encontró que también eran proteínas: pepsina, tripsina, quimotripsina, carboxipeptidasa y la enzima Old Yellow (una flavoproteína NADPH oxidasa). Desde entonces se ha demostrado que casi todas las enzimas son proteínas, o proteínas más
cofactores. Algunas moléculas de ARN también presentan actividad catalítica, pero usualmente no se les llama enzimas.
Se ha tenido la ocasión de comprobar de qué manera las formas tridimensionales de las proteínas les permiten desempeñar papeles estructurales y de transporte. Ahora se describirán sus funciones como enzimas. Las enzimas son catalizadores biológicos selectivos de una eficiencia extraordinaria. Toda célula viva dispone de cientos de enzimas distintas que catalizan las reacciones esenciales para la vida. Aun los organismos vivos más simples contienen múltiples copias de cientos de enzimas diferentes.
Las enzimas son muy específicas para los reactivos o sustratos sobre los que actúan, y varía el grado de especificidad hacia el sustrato. Algunas enzimas actúan sobre un grupo de sustratos relacionados y otras sólo sobre un simple compuesto. Muchas enzimas poseen estereoespecificidad ya que sólo actúan sobre un estereoisómero del sustrato. Quizá el aspecto más importante de la especificidad de una enzima es la especificidad de reacción, esto es, la falta de formación de subproductos como desperdicios. La especificidad de reacción se refleja en la pureza excepcional del producto (100% en esencia) —mucho mayor que la pureza de productos de reacciones típicas catalizadas en química orgánica.
Las enzimas pueden hacer más que sólo aumentar la velocidad de una sola reacción muy específica. Algunas también pueden combinar, o acoplar, dos reacciones que normalmente serían separadas. Esta propiedad permite que la energía ganada en una reacción se use en una segunda reacción. Las reacciones acopladas son una propiedad común de muchas enzimas; por ejemplo, la hidrólisis del ATP se acopla con frecuencia a reacciones metabólicas menos favorables.

Las seis clases de enzimas
Los nombres en la mayor parte de las enzimas metabólicas se forman agregando el sufijo—asa al nombre de sus sustratos, o a un término descriptivo de la reacción que catalizan. Por ejemplo, la ureasa tiene a la urea como sustrato.
1. Las oxidorreductasas catalizan las reacciones de oxidación-reducción. La mayor parte de esas enzimas se llaman, en general, deshidrogenasas. También hay otras enzimas en esta clase que se llaman oxidasas, peroxidasas, oxigenasas o reductasas. En bioquímica hay cada vez más la tendencia a citar esas enzimas por su nombre formal, oxidorreductasas, y no por los nombres más comunes en las publicaciones no muy recientes de bioquímica. Un ejemplo de una oxidorreductasa es la lactato deshidrogenasa (EC 1.1.1.27), llamada también lactato: NAD oxidorreductasa. Esta enzima cataliza la conversión reversible de L-lactato en piruvato. La oxidación de L-lactato se acopla a la reducción de la coenzima nicotinamida adenina dinucleótido (NAD_).

2. Las transferasas catalizan las reacciones de transferencia de un grupo y pueden necesitar la presencia de coenzimas. En las reacciones de transferencia de grupo, una parte de la molécula del sustrato se suele enlazar en forma covalente con la enzima o con su coenzima. Este grupo incluye las cinasas, enzimas que catalizan la transferencia de un grupo fosforilo del ATP. La alanina transaminasa, cuyo nombre sistemático es L-alanina:2-oxiglutarato aminotransferasa (EC 2.6.1.2), es un ejemplo típico de esta clase.

3. Las hidrolasas catalizan hidrólisis. Son una clase especial de transferasas donde el agua sirve como aceptor del grupo transferido. La pirofosfatasa es un ejemplo sencillo de una hidrolasa. El nombre sistemático de esta enzima es difosfato fosfohidrolasa (EC 3.6.1.1).

4. Las liasas catalizan la lisis de un sustrato, al generar un enlace doble; son reacciones de eliminación, no hidrolíticas y no oxidantes. En dirección inversa, las liasas catalizan la adición de un sustrato a un doble enlace de un segundo sustrato. Una liasa que cataliza una reacción de adición en las células es frecuentemente llamada sintasa. La piruvato descarboxilasa pertenece a esta clase de enzimas ya que descompone al piruvato en acetaldehído y dióxido de carbono. El nombre sistemático de la piruvato descarboxilasa, 2-oxo-ácido carboxi-liasa (EC 4.1.1.1.) casi nunca se emplea.

5. Las Isomerasas catalizan cambios estructurales dentro de una misma molécula (reacciones de isomerización). Como estas reacciones sólo tienen un sustrato y un producto son de las reacciones enzimáticas más simples. La alanina racemasa (EC 5.1.1.1) es una isomerasa que cataliza la interconversión de L-alanina y D-alanina. El nombre común es igual al nombre sistemático.

6. Las ligasas catalizan la ligadura o unión de dos sustratos. Estas reacciones necesitan un suministro de energía potencial química de un nucleósido trifosfato, como el ATP. Las ligasas son usualmente llamadas sintetasas. La glutamina sintetasa, o L-glutamato: amoniaco ligasa (formadora de ADP) (EC 6.3.12) usa la energía de la hidrólisis del ATP para unir glutamato y amoniaco para producir glutamina.

Inhibición reversible de enzimas
Un inhibidor de enzima (I) es un compuesto que se enlaza con una enzima e interfiere con su actividad. Los inhibidores pueden actuar evitando la formación del complejo ES o bloqueando la reacción química que lleva a la formación del producto. Por regla general, los inhibidores son moléculas pequeñas que se unen en forma reversible con la enzima que inhiben. Las células contienen muchos inhibidores enzimáticos naturales que juegan papeles importantes en la regulación del metabolismo. Los inhibidores artificiales se usan en experimentos para investigar los mecanismos enzimáticos y para descifrar las rutas metabólicas. Algunas medicinas y muchos venenos son inhibidores de enzimas. Algunos inhibidores se unen en forma covalente con las enzimas y causando que la inhibición sea irreversible. La mayor parte de la inhibición de relevancia biológica es reversible. Los inhibidores reversibles se unen a las enzimas con las mismas fuerzas no covalentes que enlazan a sustratos y productos. Los inhibidores reversibles se diferencian de los irreversibles por su fácil eliminación de soluciones de enzima por métodos como diálisis o filtración en gel. El equilibrio entre la enzima libre (E) más el inhibidor (I) y el complejo EI se caracteriza por una constante de disociación. En este caso, a la constante se le llama constante de inhibición, Ki.
Inhibición competitiva
Los inhibidores competitivos son los que se encuentran con más frecuencia en bioquímica. En la inhibición competitiva, el inhibidor sólo se puede unir a moléculas de enzima libre que no estén unidas a sustrato alguno. La formación de un complejo EI quita a la enzima de su ruta normal. Cuando un inhibidor competitivo se une con una molécula de enzima, una molécula de sustrato no puede unirse a esa molécula de enzima. Al revés, la unión de sustrato y una molécula de enzima evitan el enlazamiento de un inhibidor. En otras palabras, S e I compiten por unirse a la molécula de enzima. Más comúnmente, S e I se unen al mismo sitio de la enzima, el sitio activo. Este tipo de inhibición se llama inhibición competitiva clásica. No es la única clase de inhibición competitiva. En algunos casos, como en las enzimas alostéricas, el inhibidor se une a un sitio diferente, lo que altera el sitio de unión del sustrato y evita esta unión. A este tipo de inhibición se le llama inhibición competitiva no clásica. Cuando están presentes tanto I como S en una solución, la proporción de la enzima que puede formar complejos IS depende de las concentraciones de sustrato e inhibidor y de sus afinidades relativas hacia la enzima.
La cantidad de EI se puede reducir aumentando la concentración de S. A concentraciones suficientemente altas, la enzima puede estar saturada con sustrato.
Muchos inhibidores competitivos clásicos son análogos al sustrato, compuestos que se parecen en su estructura a los sustratos. Los análogos se unen a la enzima, pero no reaccionan. Por ejemplo, la benzamidina es un inhibidor competitivo de la tripsina. La tripsina cataliza la hidrólisis de los enlaces peptídicos cuyos grupos carbonilo contienen residuos de arginina y lisina, y la benzamidina es un análogo de la cadena lateral de alquilguanidilo en la arginina. La benzamidina actúa como inhibidor que compite con los residuos de arginina en péptidos por la unión de la tripsina.

Inhibición acompetitiva
Los inhibidores acompetitivos sólo se unen al ES y no a la enzima libre.
En la inhibición acompetitiva disminuye la Vmáx (aumenta 1/Vmáx) por conversión de algunas moléculas de E en la forma inactiva ESI. Ya que es el complejo ES el que se enlaza con I y la disminución de Vmáx no se revierte por la adición de más sustrato.
También, los inhibidores acompetitivos hacen descender la Km (vista como un aumento del valor absoluto de 1/Km en una gráfica de doble recíproco) ya que los equilibrios de formación de ES y de ESI son desplazados hacia los complejos, por la unión de I. Experimentalmente, las líneas de una gráfica de doble recíproco representando concentraciones variables de un inhibidor acompetitivo, tienen todas la misma pendiente, lo que indica que los valores de Km y de Vmáx decrecieron proporcionalmente. Este tipo de inhibición suele presentarse en las reacciones de multisustrato.

Inhibición no competitiva
Los inhibidores no competitivos se pueden unir a la E o al ES y formar complejos inactivos EI o ESI, respectivamente. Esos inhibidores no son análogos del sustrato y no se enlazan en el mismo sitio que el S. El caso clásico de inhibición no competitiva se caracteriza por una disminución aparente de Vmáx (1/Vmáx parece aumentar) sin cambiar de la Km. En una gráfica de doble recíproco, las líneas de la inhibición no competitiva clásica se cruzan en el punto del eje x que corresponde a –1/Km. Esta ordenada al origen común indica que Km no se afecta. El efecto de la inhibición no competitiva está dado por la interacción del I con la E y el ES en forma reversible, eliminando las moléculas de enzima activa en la solución. Esta inhibición no se puede compensar agregando S. Es rara la inhibición no competitiva clásica, pero se conocen ejemplos entre las enzimas alostéricas. En esos casos, es probable que el inhibidor no competitivo altere la conformación de la enzima, cuya forma todavía le permita seguir uniéndose al S pero sin poder catalizar reacción alguna. La mayor parte de las enzimas no se apega a la forma clásica de inhibición no competitiva, donde no cambia Km. En la mayoría de los casos se afectan tanto la Vmáx como la Km, ya que la afinidad del inhibidor hacia la E es distinta que hacía ES. En esos casos se suelen llamar de inhibición mixta.

Inhibición enzimática irreversible
En contraste con un inhibidor enzimático reversible, un inhibidor enzimático irreversible forma un enlace covalente estable con una molécula de enzima y elimina así las moléculas del sitio activo en la población enzimática. Típicamente, la inhibición irreversible ocurre por alquilación o acilación de la cadena lateral de un residuo de aminoácido en el sitio activo. Hay muchos inhibidores irreversibles naturales, al igual que hay ejemplos sintéticos que se describirán aquí. Una aplicación importante de los inhibidores irreversibles es la identificación de residuos de aminoácidos en el sitio activo, por sustitución específica de sus cadenas laterales reactivas. En este proceso, un inhibidor irreversible que sólo reacciona con un tipo de aminoácido se incuba con una solución de la enzima, la que a continuación es analizada para determinar su pérdida de actividad. Las cadenas laterales ionizables se modifican con reacciones de acilación o alquilación. Por ejemplo, los grupos amino libres, como el grupo e-amino de la lisina, reaccionan con un aldehído para formar una base de Schiff, la cual se puede estabilizar por reducción con borohidruro de sodio (NaBH4).

Carbohidrato
 Los carbohidratos (también llamados “hidratos de carbono”) son uno de los tres tipos de macronutrientes presentes en nuestra alimentación (los otros dos son las grasas y las proteínas). Existen en multitud de formas y se encuentran principalmente en los alimentos tipo almidón, como el pan, la pasta alimenticia y el arroz, así como en algunas bebidas, como los zumos de frutas y las bebidas endulzadas con azúcares. Los carbohidratos constituyen la fuente energética más importante del organismo y resultan imprescindibles para una alimentación variada y equilibrada.
Los carbohidratos (también llamados “hidratos de carbono”) son uno de los tres tipos de macronutrientes presentes en nuestra alimentación (los otros dos son las grasas y las proteínas). Existen en multitud de formas y se encuentran principalmente en los alimentos tipo almidón, como el pan, la pasta alimenticia y el arroz, así como en algunas bebidas, como los zumos de frutas y las bebidas endulzadas con azúcares. Los carbohidratos constituyen la fuente energética más importante del organismo y resultan imprescindibles para una alimentación variada y equilibrada.
El progreso en las investigaciones científicas ha puesto en relieve las diversas funciones que tienen los carbohidratos en el cuerpo y su importancia para gozar de una buena salud. En la siguiente explicación se examinan más a fondo dichas investigaciones, para que el lector conozca mejor este macronutriente, siendo además necesario señalar que gran parte de nuestros conocimientos en torno a los carbohidratos datan ya de hace bastante tiempo.
¿Qué son los carbohidratos?
Todos los carbohidratos están formados por unidades estructurales de azúcares, que se pueden clasificar según el número de unidades de azúcar que se combinen en una molécula. La glucosa, la fructosa y la galactosa son ejemplos destacados de los azúcares constituidos por una sola unidad (de azúcar); dicho tipo de azúcares se conocen también como “monosacáridos”. A los azúcares constituidos por dos unidades se le denomina “disacáridos”; los disacáridos más ampliamente conocidos son la sacarosa (“azúcar de mesa”) y la lactosa (el azúcar de la leche). La tabla siguiente muestra los principales tipos de carbohidratos alimenticios.
CLASIFICACIÓN DE LOS CARBOHIDRATOS ALIMENTICIOS y ejemplos correspondientes
CLASE
|
EJEMPLOS
|
Monosacáridos
|
Glucosa, fructosa, galactosa
|
Disacáridos
|
Sacarosa, lactosa, maltosa
|
Polioles
|
Isomaltol, maltitol, sorbitol, xilitol, eritritol
|
Oligosacáridos
|
Fructooligosacáridos, maltooligosacáridos
|
Polisacáridos tipo almidón
|
Amilosa, amilopectina, maltodextrinas
|
Polisacáridos no semejantes al almidón (fibra alimenticia)
|
Celulosa, pectinas, hemicelulosas, gomas, inulina
|
Azúcares
La glucosa y la fructosa son monosacáridos y se pueden encontrar en las frutas, las vallas, las verduras, la miel y los siropes de glucosa-fructosa. El azúcar común o de mesa, es decir, la sacarosa, es un disacárido compuesto por glucosa y fructosa y está presente en la naturaleza en alimentos tales como la remolacha azucarera, la caña de azúcar y las frutas. La lactosa, que es un disacárido compuesto de glucosa y galactosa, es el principal azúcar de la leche y de los productos lácteos; por su parte, la maltosa, que es un disacárido compuesto sólo de glucosa (dos moléculas de glucosa), está presente en la malta y en los siropes (extractos líquidos) derivados del almidón. Tanto el azúcar de mesa (sacarosa) y los siropes de glucosa-fructosa contienen glucosa y fructosa, bien en estado libre (siropes de glucosa-fructosa) o en forma de disacárido (sacarosa).

Los polioles se denominan alcoholes de azúcar. Hay polioles naturales, pero la mayoría se fabrican mediante la transformación de azúcares. El poliol utilizado con mayor frecuencia es el sorbitol; por su parte, el xilitol se usa frecuentemente en las gomas de mascar y en los caramelos. El isomaltol es otro poliol, que se usa en repostería/confitería y se obtiene a partir de la sacarosa. Los polioles son dulces y se pueden utilizar en los alimentos (añadiéndolos a los mismos) de forma similar a lo que se hace con los azúcares, aunque dichos polioles pueden tener un efecto laxante si se ingieren en cantidades excesivas.
Oligosacáridos
La Organización Mundial de la Salud (OMS) define a los oligosacáridos como carbohidratos formados por 3-9 unidades de azúcares (monosacáridos), aunque en otras definiciones se habla de cadenas de azúcares ligeramente más largas. Los fructooligosacáridos contienen un total de hasta 9 unidades de fructosa y se producen con fines comerciales mediante la hidrólisis (descomposición enzimática) parcial de la inulina. La rafinosa y la estaquiosa están presentes, si bien en cantidades pequeñas, en determinadas legumbres, cereales y verduras, así como en la miel.

Polisacáridos
Se necesitan más de 10 unidades de azúcar y a veces hasta miles de unidades para formar los polisacáridos. El almidón es la principal reserva de energía de las hortalizas de raíz y los cereales. Está formado por largas cadenas de glucosa en forma de gránulos, cuyo tamaño y forma varían según el vegetal del que forma parte. El equivalente de los almidones en los animales y en los seres humanos es el llamado “glucógeno” (ver sección 3.1).

Los polisacáridos sin almidón son los principales componentes de la fibra alimenticia. Comprenden: celulosa, hemicelulosa, inulina, pectinas y gomas. La celulosa es el componente principal de las paredes celulares vegetales y está formada por miles de unidades de glucosa. Los distintos componentes de la fibra alimenticia tienen diferentes propiedades y estructuras físicas. Una característica distintiva de la fibra alimenticia es que no puede ser digerida por los seres humanos. Sin embargo, algunos tipos de fibra pueden ser metabolizados por las bacterias intestinales, dando lugar a compuestos que las células intestinales humanas sí que pueden utilizar para la producción de energía. En cualquier caso, por no poder ser digerida por los seres humanos, la fibra tiene un menor contenido energético medio que la mayoría de los demás carbohidratos (ver sección 3.1).
Los carbohidratos en el cuerpo
La función principal de los carbohidratos es proporcionar energía, aunque también desempeñan una función importante para la estructura y el funcionamiento de las células, tejidos y órganos; además, sirven para formar las estructuras carbohidratadas de la superficie de las células. Hay diversas clases de moléculas carbohidratadas en el cuerpo: proteoglicanos, glucoproteínas (también llamadas “glicoproteínas”), y glucolípidos (también llamados “glicolípidos”).
Fuente y almacenamiento de energía
Los almidones y los azúcares son las principales fuentes de energía y aportan 4 kilocalorías (17 kilojulios) por gramo. Los polioles proporcionan 2,4 kilocalorías (10 kilojulios), y la fibra alimenticia, 2 kilocalorías (8 kilojulios) por gramo, respectivamente. Nota importante: el poliol eritritol no es metabolizado en absoluto por el cuerpo y, por eso, proporciona cero calorías.
En el intestino delgado, los monosacáridos son absorbidos y de allí pasan al torrente sanguíneo, desde donde son transportados hasta los lugares en los que son utilizados. Los disacáridos son descompuestos en azúcares simples por las enzimas digestivas. El cuerpo también necesita la ayuda de las enzimas digestivas para romper las largas cadenas de almidones y descomponerlas en los azúcares por los que están formadas, que pasan posteriormente a la sangre.
El cuerpo humano utiliza los carbohidratos en forma de glucosa. La glucosa también se puede transformar en glucógeno, un polisacárido similar al almidón, que es almacenado en el hígado y en los músculos como fuente de energía de la que el cuerpo puede disponer fácilmente. El cerebro y los eritrocitos (“glóbulos rojos”) necesitan la glucosa, ya que no pueden emplear otra cosa como fuente de energía: ni grasas, ni proteínas, ni ninguna otra forma de energía. Por este motivo se debe mantener constantemente el nivel de glucosa en sangre en un nivel óptimo. Para cubrir las necesidades energéticas del cerebro se necesitan aproximadamente 130 gr de glucosa al día. La glucosa puede proceder directamente de los carbohidratos ingeridos con la dieta, de los depósitos de glucógeno o de la conversión de determinados aminoácidos derivados de la degradación de las proteínas. Varias hormonas, entre ellas la insulina, trabajan rápidamente para regular el flujo de glucosa que entra y sale de la sangre y mantenerla a un nivel estable.
Lípidos
Se sabe que bajo el nombre de lípidos se agrupa una serie de sustancias que tienen en común ciertas características de solubilidad en solventes orgánicos.
Dentro de este grupo heterogéneo, que genéricamente se designa por lípidos, se encuentran las materias grasas tanto sólidas como líquidas que normalmente y diariamente se ingieren junto con la dieta. Debe eso sí diferenciarse entre grasa de depósito, constituida principalmente por triglicéridos y materias grasas estructurales que, además de estos componentes, están constituidas en parte importante por fosfolípidos u otro tipo de estructuras más complejas como esfingolípidos, cerebrósidos, etc.





![]()

![]()

![]()
![]()
![]()
![]() Grupos fosfato
Grupos fosfato
Son los que dan la característica ácida al ADN y ARN. Estos ácidos nucleicos, al tener nucleótidos con un solo radical (monofosfato) son estables. Cuando el nucleótido contiene más grupos fosfato (difosfato, trifosfato) se vuelve inestable, como sucede con el adenosin trifosfato o ATP. En consecuencia, se rompe un enlace fosfato y se libera la energía que lo une al nucleótido. Los grupos fosfato forman parte de la bicapa lipídica de las membranas celulares.
![]()
Pentosas
Son monosacáridos con cinco carbono en su molécula. En los ácidos nucleicos hay dos tipos de pentosas, la desoxirribosa presente en el ADN y la ribosa, que forma parte del ARN.
Bases nitrogenadas
También hay dos tipos. Las derivadas de la purina son la adenina y la guanina y las que derivan de la pirimidina son la citosina, la timina y el uracilo.
![]()

![]()


Los lípidos al igual que la proteínas y los carbohidratos son componentes esenciales de todos los organismos vivos.
Los lípidos son moléculas bastantes pequeñas que presentan una fuerte tendencia a asociarse mediante fuerzas no covalentes.
Son compuestos constituidos al igual que los carbohidratos de C, H y O; sin embargo la relación de H:O de los lípidos es mucho menor que la relación de los carbohidratos.
Se sabe que bajo el nombre de lípidos se agrupa una serie de sustancias que tienen en común ciertas características de solubilidad en solventes orgánicos.
Dentro de este grupo heterogéneo, que genéricamente se designa por lípidos, se encuentran las materias grasas tanto sólidas como líquidas que normalmente y diariamente se ingieren junto con la dieta. Debe eso sí diferenciarse entre grasa de depósito, constituida principalmente por triglicéridos y materias grasas estructurales que, además de estos componentes, están constituidas en parte importante por fosfolípidos u otro tipo de estructuras más complejas como esfingolípidos, cerebrósidos, etc.
Los ácidos grasos son los principales constituyentes de los lípidos neutros (triglicéridos) y lípidos polares (fosfolípidos, esfingolípidos, etc.) ya que se encuentran esterificando un alcohol el cual se puede considerar como un soporte. Por este motivo, se presentarán en este trabajo los principales ácidos grasos constituyentes de diferentes materias grasas de origen vegetal, animal, marino, e hidrogenadas y su respectiva distribución porcentual.
· Función de los lípidos en el cuerpo humano
Ø Fuente y Reserva de energía
§ Los triacilgliceroles, son las moléculas intracelulares de almacenamiento de energía metabólica.
§ La mayor parte de la grasa de los animales se oxida para generar energía en forma de ATP e impulsa los procesos metabólicos.
§ Un gramo de grasa contiene aproximadamente 9kcal de energía.
§ Los adultos con un porcentaje corporal de 10 a 30% la reserva de energía disponible oscila entre 70 000 a 110000 kcal aproximadamente.
Ø Función estructural: Forman las bicapas lipídicas de las membranas. Recubren órganos y le dan consistencia, o protegen mecánicamente como el tejido adiposo de pies y manos.
Ø Función biocatalizadora: En este papel los lípidos favorecen o facilitan las reacciones químicas que se producen en los seres vivos. Cumplen esta función las vitaminas lipídicas, las hormonas esteroideas y las prostaglandinas.
Ø Función transportadora: El transporte de lípidos desde el intestino hasta su lugar de destino se realiza mediante su emulsión gracias a los ácidos biliares y a los proteolípidos.
Transporte de acido graso activado a través de la membrana mitocondrial. El acido graso activado es transportado al espacio intra-membranal mediante la carnitina aciltransferasa I. Una vez ahí, es llevada a la matriz mediante difusión facilitada donde se separa de la carnitina. La carnitina puede retornar inmediatamente al citoplasma.

· Diversidad estructural y función de los lípidos

· Glucolípidos
Los glucolípidos forman parte de los carbohidratos de la membrana celular, que están unidos a lípidos únicamente en el exterior de la membrana plasmática y en el interior de algunos organelos.
Entre los principales glúcidos que forman los glucolípidos
encontramos a la galactosa, manosa, fucosa, glucosa, glucosamina,
galactosamina y el ácido siálico. Entre los glicolípidos más comunes
están los cerebrósidos y gangliósidos.

· Clasificación de los lípidos
Lípidos simples
Los lípidos simples son aquellos que solo consisten un acido graso en su estructura. Dentro de este grupo de lípidos tenemos a :
• Ácidos grasos
Los ácidos grasos consiste en una cadena hidrocarbonada lineal,
cosí siempre con numero par de átomos de carbono.
Reciben su nombre debido a la molécula de acido orgánico (COOH), que forma parte de la estructura química.
Los ácidos grasos se almacenan en forma de triacilglicerol.
Los ácidos grasos varían en su longitud, nivel de saturación.
Mientras que los ácidos grasos saturados solo tienen enlaces simples entre los átomos de carbono y contienen el mayor número posible de átomos de hidrógeno en la cadena de carbono, los ácidos grasos insaturados tienen enlaces dobles o triples en su cadena hidrocarbonada y son aquellos que han perdido algunos átomos de hidrógeno.

• Triglicéridos
Son combustibles metabólicos importantes, en especial en los mamíferos.
La oxidación de los ácidos grasos produce mas energía (37 kJ g-1) que la oxidación de proteínas y carbohidratos (16 kJ g-1 cada uno).
Los ácidos grasos se almacenan en forma de lípidos neutros llamados triacilgliceroles.
Los triacilgliceroles están formados por tres residuos de acilo graso esterificados con glicerina.
Los triacilgliceroles son muy hidrofóbicos. Se almacenan en células en forma anhidra, las moléculas no están solvatadas por el agua.
Las grasas y los aceites son mezclas de triacilgliceroles.
Los triglicéridos son un tipo de grasa presente en el torrente sanguíneo y en el tejido adiposo. Un exceso en este tipo de grasa puede contribuir al endurecimiento y el estrechamiento de las arterias. Eso lo pone en riesgo de tener un infarto o un ataque cerebral (derrame). Enfermedades como la diabetes, la obesidad, la insuficiencia renal o el alcoholismo pueden causar un aumento de los triglicéridos.

Lípidos complejos
Los lípidos complejos son aquellos que consisten en moléculas de lípidos unos a otros compuestos químicos. Dentro de este grupo de lípidos tenemos a :
• Fosfolípidos
Los fosfolípidos en general son aquellos lípidos que contienen ácido fosfórico. En el campo de la ciencia y la tecnología de los alimentos, la expresión suele limitarse a los derivados del ácido glicerofosfórico, que están formados por una molécula de glicerol esterificada en las posiciones 1 y 2 por dos ácidos grasos, con la posición 3 esterificada por un ácido fosfórico que lleva unidas además otras estructuras, dependiendo del fosfolípido de que se trate. De forma genérica se denominan "lecitinas", aunque se considera que la lecitina propiamente dicha es la fosfatidilcolina.
Los fosfolípidos son los principales cosntituyentes lipídicos de las membranas biológicas, donde forman estructuras en bicapa, con las zonas no polares de los constituyentes de cada capa orientadas hacia el interior. Consecuentemente, los fosfolípidos se van a encontrar presentes en la mayoría de los alimentos complejos, en los que exista material celular. Los fosfolípidos son también capaces de producir estructuras artificiales del tipo de bicapa (liposomas).

• Glucolipidos
Los glucolípidos son biomoléculas compuestas por un lípido y un grupo glucídico o hidrato de carbono.
Los glucolípidos forman parte de los carbohidratos de la membrana celular, que están unidos a lípidos únicamente en el exterior de la membrana plasmática y en el interior de algunos organelos. Entre los principales glúcidos que forman los glucolípidos encontramos a lagalactosa, manosa, fucosa, glucosa, glucosamina, galactosamina y el ácido siálico. Entre los glucolípidos más comunes están los cerebrósidos y gangliósidos.
Los glucolípidos forman parte de los carbohidratos de la membrana celular, que están unidos a lípidos únicamente en el exterior de la membrana plasmática y en el interior de algunos organelos. Entre los principales glúcidos que forman los glucolípidos encontramos a lagalactosa, manosa, fucosa, glucosa, glucosamina, galactosamina y el ácido siálico. Entre los glucolípidos más comunes están los cerebrósidos y gangliósidos.

• Lipoproteínas
Aunque el término lipoproteína podría describir cualquier asociación dec lípidos con proteínas, se suele restringir para un grupo concreto de complejos moleculares que se encuentran en el plasma sanguíneo de los mamíferos; las lipoproteínas están formadas por lípidos asociados de forma no covalente con proteínas (apolipoproteínas o apoproteínas), pero también incluyen moléculas antioxidantes liposolubles. Son partículas con un centro apolar que incluye triacilgliceroles y ésteres de colesterol y un revestimiento anfifílico formado por fosfolípidos, colesterol no esterificado y las apoproteínas.
La función de las lipoproteínas plasmáticas es transportar moléculas lipídicas de unos órganos a otros en el medio acuoso del plasma. En el estado de ayuno normal el plasma humano tiene cuatro clases de lipoproteínas y en el periodo postabsortivo aparece una quinta clase, los quilomicrones.
Las lipoproteínas se clasifican en función de su densidad:
- Los quilomicrones son lipoproteínas grandes con densidad extremadamente baja que transportan los lípidos de la dieta desde el intestino a los tejidos.
- Las VLDL, lipoproteínas de muy baja densidad, se sintetizan en el hígado y transportan lípidos a los tejidos; estas VLDL van perdiendo en el organismo triacilgliceroles y algunas apoproteínas y fosfolípidos; finalmente sus restos sin triacilgliceroles (IDL, lipoproteínas de densidad intermedia) son captados por el hígado o convertidos en LDL.
- Las LDL, lipoproteínas de baja densidad, transportan colesterol a los tejidos donde hay receptores de LDL.
- Las HDL, lipoproteínas de alta densidad, también se producen en el hígado y eliminan de las células el exceso de colesterol llevándolo al hígado, único órgano que puede desprenderse de éste convirtiéndolo en ácidos biliares.

· Colesterol
Pese a tener consecuencias perjudiciales en altas concentraciones, es esencial para crear la membrana plasmática que regula la entrada y salida de sustancias que atraviesan la célula.
Abundan en las grasas de origen animal.
El colesterol es un esterol (lípido) que se encuentra en los tejidos corporales y en el plasma sanguíneo de los vertebrados. Se presenta en altas concentraciones en el hígado, médula espinal, páncreas y cerebro.
Es la tercera clase de lípidos (Colesterol) que se encuentran en las membranas de los eucariotas y muy rara vez en bacterias.
Los esteroides junto con las vitaminas lipídicas y los terpenos, se clasifican como isoprenoides, por que su molécula se relaciona con le isopreno.
Los esteroides contiene cuatro anillos fundidos, tres de seis carbonos idénticos como A, B y C y un anillo D de cinco carbonos.
La estructura anular característica se deriva del ESCUALENO

ACIDOS NUCLEICOS
Son compuestos orgánicos de elevado peso molecular, formados por carbono, hidrógeno, oxígeno, nitrógeno y fósforo. Cumplen la importante función de sintetizar las proteínas específicas de las células y de almacenar, duplicar y transmitir los caracteres hereditarios. Los ácidos nucleicos, representados por el ADN (ácido desoxirribonucleico) y por el ARN (ácido ribonucleico), son macromoléculas formadas por la unión de moléculas más pequeñas llamadas nucleótidos.
· NUCLEÓTIDOS
Son moléculas compuestas por grupos fosfato, un monosacárido de cinco carbonos (pentosa) y una base nitrogenada. Además de constituir los ácidos nucleicos forman parte de coenzimas y de moléculas que contienen energía. Los nucleótidos tienen importantes funciones, entre ellas el transporte de átomos en la cadena respiratoria mitocondrial, intervenir en el proceso de fotosíntesis, transporte de energía principalmente en forma de adenosin trifosfato (ATP) y transmisión de los caracteres hereditarios.
Esquema de un nucleótido
 Grupos fosfato
Grupos fosfatoSon los que dan la característica ácida al ADN y ARN. Estos ácidos nucleicos, al tener nucleótidos con un solo radical (monofosfato) son estables. Cuando el nucleótido contiene más grupos fosfato (difosfato, trifosfato) se vuelve inestable, como sucede con el adenosin trifosfato o ATP. En consecuencia, se rompe un enlace fosfato y se libera la energía que lo une al nucleótido. Los grupos fosfato forman parte de la bicapa lipídica de las membranas celulares.

Pentosas
Son monosacáridos con cinco carbono en su molécula. En los ácidos nucleicos hay dos tipos de pentosas, la desoxirribosa presente en el ADN y la ribosa, que forma parte del ARN.
Bases nitrogenadas
También hay dos tipos. Las derivadas de la purina son la adenina y la guanina y las que derivan de la pirimidina son la citosina, la timina y el uracilo.
Bases nitrogenadas

La timina está presente solo en el ADN, mientras que el uracilo está únicamente en el ARN. El resto de las bases nitrogenadas forma parte de ambos ácidos nucleicos.
![]() La asociación de los nucleótidos con otras estructuras moleculares permite la transmisión de caracteres hereditarios y el transporte de energía.
La asociación de los nucleótidos con otras estructuras moleculares permite la transmisión de caracteres hereditarios y el transporte de energía.
 La asociación de los nucleótidos con otras estructuras moleculares permite la transmisión de caracteres hereditarios y el transporte de energía.
La asociación de los nucleótidos con otras estructuras moleculares permite la transmisión de caracteres hereditarios y el transporte de energía.
· NUCLEÓSIDOS
Es la unión de una pentosa con una base nitrogenada, a través del carbono 1’ del monosacárido con un nitrógeno de la base. Al establecerse la unión química se desprende una molécula de agua.
Esquema de un nucleósido

Los nucleósidos se identifican de acuerdo a la base nitrogenada de la cual provienen. Si derivan de bases purínicas llevan el sufijo “osina”. Si lo hacen de bases pirimidínicas se agrega la terminación “idina”. Además, si el nucleósido está unido a la desoxirribosa se le agrega el prefijo “desoxi”.
Nomenclatura de los nucleósidos

De acuerdo a lo señalado, un nucleótido está formado por un nucleósido unido a uno o más grupos fosfato. Los nucleótidos se identifican de manera similar que los nucleósidos, omitiendo la última vocal y añadiendo la palabra “fosfato”, por ejemplo, adenosin fosfato, desoxicitidin fosfato, uridin fosfato, etc.

Los ácidos nucleicos son larguísimas cadenas formadas por millones de nucleótidos que se unen entre sí por enlaces de fosfatos. La base nitrogenada del nucleótido se une al carbono 1’ de la molécula de pentosa y el grupo fosfato al carbono 5’. La columna vertebral de la cadena o hilera la constituyen el grupo fosfato y la pentosa.

· ACIDO DESOXIRRIBONUCLEICO (ADN)
Es una molécula sumamente compleja que contiene toda la información genética del individuo. El ADN regula el control metabólico de todas las células.
El ADN posee una doble cadena o hilera de polinucleótidos, ambas con forma helicoidal y ensamblada a manera de escalera. Es un ácido nucleico presente en el núcleo, en las mitocondrias y en los cloroplastos de todas las células eucariotas. Se dispone de manera lineal, aunque en las procariotas tiene forma circular y está disperso en el citoplasma.
Para su estudio se lo divide en cuatro estructuras.
Estructura primaria del ADN
Como fue señalado, cada nucleótido está compuesto por una molécula de ácido fosfórico, una desoxirribosa como pentosa y cuatro bases nitrogenadas que son la adenina, citosina, guanina y timina.
Estructura secundaria del ADN
El ADN está formado por dos hileras o cadenas de polinucleótidos. El nucleótido de cada hilera sigue a otro nucleótido, y este a su vez al siguiente. De esta forma, cada nucleótido se denomina de acuerdo a la secuencia de cada base nitrogenada. Por ejemplo, una de las secuencias puede ser G-T-A-C-A-T-G-C. Una determinada secuencia de nucleótidos del ADN se denomina gen. Los genes se ubican en un determinado lugar de los cromosomas, y ejercen funciones específicas.
![]()
El ADN posee una doble cadena o hilera de polinucleótidos, ambas con forma helicoidal y ensamblada a manera de escalera. Es un ácido nucleico presente en el núcleo, en las mitocondrias y en los cloroplastos de todas las células eucariotas. Se dispone de manera lineal, aunque en las procariotas tiene forma circular y está disperso en el citoplasma.
Para su estudio se lo divide en cuatro estructuras.
Estructura primaria del ADN
Como fue señalado, cada nucleótido está compuesto por una molécula de ácido fosfórico, una desoxirribosa como pentosa y cuatro bases nitrogenadas que son la adenina, citosina, guanina y timina.
Estructura secundaria del ADN
El ADN está formado por dos hileras o cadenas de polinucleótidos. El nucleótido de cada hilera sigue a otro nucleótido, y este a su vez al siguiente. De esta forma, cada nucleótido se denomina de acuerdo a la secuencia de cada base nitrogenada. Por ejemplo, una de las secuencias puede ser G-T-A-C-A-T-G-C. Una determinada secuencia de nucleótidos del ADN se denomina gen. Los genes se ubican en un determinado lugar de los cromosomas, y ejercen funciones específicas.

Las bases nitrogenadas de una cadena o hilera están orientadas hacia las bases nitrogenadas de la otra hilera complementaria, unidas entre sí por puentes de hidrógeno.
![]()

Las bases enfrentadas de cada hilera no lo hacen al azar, sino que la adenina se une siempre a la timina (A-T) mediante dos puentes de hidrógeno y la citosina hace lo propio con la guanina (C-G) a través de tres puentes de hidrógeno, tal como puede verse en el siguiente esquema. De esta forma, las dos hileras permanecen conectadas en toda su longitud.
![]()

La forma en que se disponen las cuatro bases nitrogenadas a lo largo de toda la cadena es la responsable de codificar la información genética de la célula, con instrucciones para controlar el desarrollo y las funciones del individuo. Numerosas proteínas como las histonas y factores de transcripción se adosan a la molécula de ADN con el fin de regular su expresión.
Estructura secundaria del ADN

· Estructura terciaria del ADN
El ADN no está libre dentro del núcleo de la célula, sino que está organizado en un complejo llamado cromatina. Se denomina cromatina a la estructura formada por ADN y proteínas histónicas y no histónicas. La cromatina está inmersa en el jugo nuclear cuando la célula está en interfase, es decir, entre dos mitosis. En esa etapa, la molécula de ADN forma largos y numerosos filamentos que se enrollan a sucesivas moléculas de proteínas especiales llamadas histonas. Esto produce que el ADN sufra una importante compactación, puesto que en cada enrollamiento el ADN da casi dos vueltas sobre cuatro pares de histonas. Esas histonas, que se reconocen como H2A, H2B, H3 y H4, forman el octámero de histonas al agruparse de a pares.


El ADN enrollado junto al octámero se denomina cromatosoma. Entre dos cromatosomas se ubica el ADN espaciador, al que está asociada otra proteína histónica llamada H1, que mantiene en posición al ADN en el octámero.

![]()

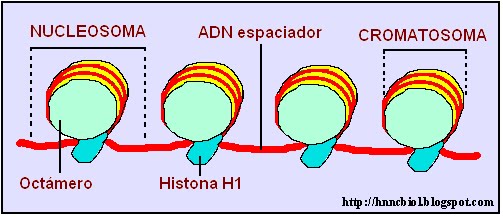
Cada cromatosoma seguido de la histona H1 y del ADN espaciador forma las unidades fundamentales de la cromatina de las células eucariotas, llamadas nucleosomas. Los nucleosomas, con unos 100 ángstrom de diámetro, adoptan la forma de un collar de perlas, forma en que se observa la cromatina mediante microscopía electrónica cuando la célula está en interfase.
Disposición en collar de perlas de la fibra de cromatina

{kind=link}
{kind=link}
{kind=link}
· Estructura cuaternaria del ADN
Los nucleosomas también se compactan enrollándose de manera helicoidal. Forman estructuras de alrededor de 300 ángstrom de diámetro, denominadas solenoides. Cuando la célula entra en mitosis, las fibras de cromatina se pliegan entre sí y se compactan aún más, formando los cromosomas.

Las proteínas no histónicas actúan como un andamiaje sobre los solenoides, ensamblándose en forma de espiral. Estas proteínas brindan un armazón a la fibra de cromatina y colaboran en su plegamiento.
![]()

· Funciones del ADN
-Almacenamiento de la información genética
-Replicación de su propia molécula
-Síntesis de ARN (transcripción)
-Transferencia de la información genética
La replicación o duplicación de la molécula de ADN se produce en la interfase de la división celular, más precisamente en la fase S, con el objetivo de conservar la información genética. Los puentes de hidrógeno que unen las dos hileras de polinucleótidos se rompen, con lo cual ambas cadenas se separan, sirviendo cada una de molde para fabricar una nueva hilera complementaria. La enzima ADN polimerasa se encarga de agregar nucleótidos fabricados por la célula que están esparcidos en el núcleo. Dicha enzima los va añadiendo a cada hilera separada conforme con la secuencia adenosina-timina y citosina-guanina (A-T y C-G). Al terminar la duplicación se obtienen dos moléculas idénticas de ADN de forma helicoidal, cada una con una hilera original y otra hilera neoformada. El núcleo tiene ahora el doble del ADN y de proteínas que al principio. De esta manera, la información genética de la célula madre será transmitida a las células hijas al producirse la mitosis.
![]()
-Replicación de su propia molécula
-Síntesis de ARN (transcripción)
-Transferencia de la información genética
La replicación o duplicación de la molécula de ADN se produce en la interfase de la división celular, más precisamente en la fase S, con el objetivo de conservar la información genética. Los puentes de hidrógeno que unen las dos hileras de polinucleótidos se rompen, con lo cual ambas cadenas se separan, sirviendo cada una de molde para fabricar una nueva hilera complementaria. La enzima ADN polimerasa se encarga de agregar nucleótidos fabricados por la célula que están esparcidos en el núcleo. Dicha enzima los va añadiendo a cada hilera separada conforme con la secuencia adenosina-timina y citosina-guanina (A-T y C-G). Al terminar la duplicación se obtienen dos moléculas idénticas de ADN de forma helicoidal, cada una con una hilera original y otra hilera neoformada. El núcleo tiene ahora el doble del ADN y de proteínas que al principio. De esta manera, la información genética de la célula madre será transmitida a las células hijas al producirse la mitosis.

· ACIDO RIBONUCLEICO (ARN)
A diferencia del ADN que posee desoxirribosa y timina, el ARN está formado por ribosa como monosacárido y uracilo como una de las bases nitrogenadas. El ARN forma una sola cadena de polinucleótidos dispuesta en manera lineal. Está presente en el citoplasma de las células procariotas y eucariotas.
La formación o síntesis de ARN se realiza a partir del ADN mediante la enzima ARN polimerasa, que copia una secuencia de nucleótidos (genes) de una hilera del ADN.
El ARN controla las etapas intermedias en la formación (síntesis) de proteínas.
Existen cuatro tipos de ARN con distintas funciones. Ellos son el ARN mensajero, el ARN de transferencia, el ARN ribosómico y el ARN heteronuclear.
-ARN mensajero (ARNm)
Se forma a partir del molde de una hilera de ADN. El ARN mensajero transporta la información para sintetizar una proteína copiada del ADN, desde el núcleo hasta el citoplasma, pasando por los poros de la membrana nuclear o carioteca. Luego se acopla a los ribosomas, organelas celulares donde se produce la síntesis de proteínas. Un codón está formado por tres nucleótidos del ARNm. Cada codón contiene un aminoácido diferente. Por lo tanto, a partir de la sucesión de los nucleótidos del ARNm se arma la secuencia de aminoácidos de la proteína. Debe recordarse que una serie de aminoácidos forman una proteína. El ARNm se degrada rápidamente por acción enzimática.
-ARN de transferencia (ARNt)
Tiene por función transportar aminoácidos hacia el ribosoma. En un extremo de su estructura, el ARNt posee un lugar específico para que se fije el aminoácido. En el otro extremo tiene un anticodón, formado por tres nucleótidos que se unen al codón del ARNm por puentes de hidrógeno.
-ARN ribosómico (ARNr)
Se unen a proteínas para formar los ribosomas, organelas formadas por dos subunidades, una mayor y otra menor. En los ribosomas se produce la síntesis de proteínas. El ARNr se sitúa en el citoplasma, y es el tipo de ácido ribonucleico más abundante de las células. El ARN nucleolar, ubicado en el nucléolo de las eucariotas, es el precursor del ARN ribosómico.
La traducción es el paso de la información transportada por el ARN-m a proteína. La especificidad funcional de los polipéptidos reside en su secuencia lineal de aminoácidos que determina su estructura primaria, secundaria y terciaria. De manera, que los aminoácidos libres que hay en el citoplasma tienen que unirse para formar los polipéptidos y la secuencia lineal de aminoácidos de un polipéptido depende de la secuencia lineal de ribonucleótidos en el ARN que a su vez está determinada por la secuencia lineal de bases nitrogenadas en el ADN.
Los elementos que intervienen en el proceso de traducción son fundamentalmente: los aminoácidos, los ARN-t (ARN transferentes), los ribosomas, ARN-r (ARN ribosómico y proteínas ribosomales), el ARN-m (ARN mensajero), enzimas, factores proteicos y nucleótidos trifosfato (ATP, GTP).
La formación o síntesis de ARN se realiza a partir del ADN mediante la enzima ARN polimerasa, que copia una secuencia de nucleótidos (genes) de una hilera del ADN.
El ARN controla las etapas intermedias en la formación (síntesis) de proteínas.
Existen cuatro tipos de ARN con distintas funciones. Ellos son el ARN mensajero, el ARN de transferencia, el ARN ribosómico y el ARN heteronuclear.
-ARN mensajero (ARNm)
Se forma a partir del molde de una hilera de ADN. El ARN mensajero transporta la información para sintetizar una proteína copiada del ADN, desde el núcleo hasta el citoplasma, pasando por los poros de la membrana nuclear o carioteca. Luego se acopla a los ribosomas, organelas celulares donde se produce la síntesis de proteínas. Un codón está formado por tres nucleótidos del ARNm. Cada codón contiene un aminoácido diferente. Por lo tanto, a partir de la sucesión de los nucleótidos del ARNm se arma la secuencia de aminoácidos de la proteína. Debe recordarse que una serie de aminoácidos forman una proteína. El ARNm se degrada rápidamente por acción enzimática.
-ARN de transferencia (ARNt)
Tiene por función transportar aminoácidos hacia el ribosoma. En un extremo de su estructura, el ARNt posee un lugar específico para que se fije el aminoácido. En el otro extremo tiene un anticodón, formado por tres nucleótidos que se unen al codón del ARNm por puentes de hidrógeno.
-ARN ribosómico (ARNr)
Se unen a proteínas para formar los ribosomas, organelas formadas por dos subunidades, una mayor y otra menor. En los ribosomas se produce la síntesis de proteínas. El ARNr se sitúa en el citoplasma, y es el tipo de ácido ribonucleico más abundante de las células. El ARN nucleolar, ubicado en el nucléolo de las eucariotas, es el precursor del ARN ribosómico.
Comentarios sobre los videos
Replicación de ADN
Una vez que se comprobó que el ADN era el material hereditario y se descifró su estructura, lo que quedaba era determinar como el ADN copiaba su información y como la misma se expresaba en el fenotipo. Matthew Meselson y Franklin W. Stahl diseñaron el experimento para determinar el método de la replicación del ADN. Tres modelos de replicación era plausibles.
Replicación conservativa durante la cual se produciría un ADN completamente nuevo durante la replicación.
En la replicación semiconservativa se originan dos moléculas de ADN, cada una de ellas compuesta de una hebra de el ADN original y de una hebra complementaria nueva. En otras palabras el ADN se forma de una hebra vieja y otra nueva. Es decir que las hebras existentes sirven de molde complementario a las nuevas.
La replicación dispersiva implicaría la ruptura de las hebras de origen durante la replicación que, de alguna manera se reordenarían en una molécula con una mezcla de fragmentos nuevos y viejos en cada hebra de ADN.
La replicación del ADN, que ocurre una sola vez en cada generación celular, necesita de muchos "ladrillos", enzimas, y una gran cantidad de energía en forma de ATP. La replicación del ADN en el ser humano a una velocidad de 50 nucleótidos por segundo, en procariotas a 500/segundo. Los nucleótidos tienen que se armados y estar disponibles en el núcleo conjuntamente con la energía para unirlos.
Transcripción de ADN A ARN
Una vez que se conforman las dos cadenas nuevas de ADN, lo que sigue es pasar la información contenida en estas cadenas a una cadena de ARN, proceso que se conoce como transcripción. Aquí la enzima responsable es la ARN polimerasa, la cual se une a una secuencia específica en el ADN denominada promotor y sintetiza ARN a partir de ADN.
ADN ---------> ARN --------> PROTEINAS replicación --> transcripción --> traducción
La síntesis de ARN usando un ADN patrón es llamada transcripción, mientras que la síntesis de proteína a partir de un ARN patrón es llamado traducción.
La secuencia de ADN transcrito por la ARN polimerasa en ARN es llamado UNIDAD DE TRANSCRIPCIÓN y el producto de la síntesis (ARN) es el TRANSCRIPTO PRIMARIO.
La transcripción se produce
Desde el codón de inicio hasta la secuencia terminadora
Desde el promotor hasta el codón de stop
Desde el codón de inicio hasta el codón de stop
Desde el promotor hasta la secuencia de terminación
Desde el codón de inicio hasta la secuencia terminadora
Desde el promotor hasta el codón de stop
Desde el codón de inicio hasta el codón de stop
Desde el promotor hasta la secuencia de terminación
Traducción (de ADN A proteína)
La traducción es el paso de la información transportada por el ARN-m a proteína. La especificidad funcional de los polipéptidos reside en su secuencia lineal de aminoácidos que determina su estructura primaria, secundaria y terciaria. De manera, que los aminoácidos libres que hay en el citoplasma tienen que unirse para formar los polipéptidos y la secuencia lineal de aminoácidos de un polipéptido depende de la secuencia lineal de ribonucleótidos en el ARN que a su vez está determinada por la secuencia lineal de bases nitrogenadas en el ADN.
Los elementos que intervienen en el proceso de traducción son fundamentalmente: los aminoácidos, los ARN-t (ARN transferentes), los ribosomas, ARN-r (ARN ribosómico y proteínas ribosomales), el ARN-m (ARN mensajero), enzimas, factores proteicos y nucleótidos trifosfato (ATP, GTP).
No hay comentarios.:
Publicar un comentario